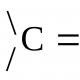Цитоскелет клітини будови. Цитоскелет. Функції цитоскелета. Будова цитоскелета. Кресцентин, гомолог білків проміжних філаментів
Еукаріотичні клітини здатні змінювати свою форму, переміщатися, пересувати органели по цитоплазмі та розділяти хромосоми під час мітозу. Ця здатність забезпечується тривимірною мережею білкових ниток (філаментів), що становлять головну архітектуру клітини – цитоскелет (іноді позначається як цитоматрікс). Білкові волокна пронизують цитоплазму еукаріотичних клітин і в безлічі точок пов'язані з білками плазматичної мембрани та органел. Всі ці волокна є структури, що складаються з субодиниць - спеціальних глобулярних білків. (Білки цитоскелета, як і інші білки клітини, закодовані в генах і синтезуються на рибосомах.)
Субодиниці цитоскелета з'єднуються між собою слабкими зв'язками (водневими, іонними та ін) і ця властивість дозволяє клітині формувати динамічні просторові структури цитоскелета, що легко змінюються. Зазначено, що з різних впливах клітина насамперед перебудовує цитоскелет, демонтуючи основні компоненти своєї архітектури, та був формує їх заново, відповідно до характером отриманого сигналу; при цьому детальна будова цитоскелета постійно змінюється за збереження загального плану його організації. Таку форму роботи цитоскелетної системи називають принципом динамічної нестабільності.
Залежно від діаметра філаменти поділяються на три групи: мікрофіламенти (5–7 нм), проміжні волокна (близько 10 нм) та мікротрубочки (близько 25 нм). Кожен тип цитоскелетних структур утворює в клітині власну систему зі своїми основними та мінорними білками. Ці системи є абсолютно незалежними, а взаємодіють друг з одним і коїться з іншими компонентами клітини – плазматичної мембраною, ядром та інші органоїдами клітини. Згідно з існуючими уявленнями, цитоскелет не тільки сприяє підтримці форми клітини та здійснює всі типи клітинних рухів, але й поєднує різні частини клітини та забезпечує передачу сигналів усередині клітини за рахунок утворення просторових білкових комплексів між рецепторами та ферментами.
Мікрофіламентизустрічаються практично у всіх типах клітин та складаються з білка актину – найбільш поширеного в еукаріотичних клітинах. (Актин становить близько 5% загального білка клітини; у скелетних м'язах – приблизно 20% клітинної маси.) Актин може існувати у вигляді мономеру (G-актин – «глобулярний актин», що складається з 375 амінокислотних залишків) або волокна (F-актин – "Фібрилярний актин"). Кожен F-актиновий філамент репрезентує спіралеподібну структуру довжиною кілька мікрометрів. Волокна F-актину має два різноіменно заряджені кінці, які полімеризуються з різною швидкістю. Швидко зростаючий кінець називається плюс-кінцем, а повільно зростаючий – мінус-кінцем. Плюс-кінець актинового філаменту росте в 10 разів швидше, ніж мінус-кінець.
Мікрофіламенти беруть участь у динамічних процесах, таких як м'язове скорочення, рух нем'язових клітин, фагоцитоз, утворення виростів цитоплазми у рухомих клітин і акросом в процесі злиття сперматозоїда з яйцеклітиною. Всі ці процеси здійснюються за допомогою актин-зв'язуючих білків.
У цитоплазмі клітин є більше 50 різних типів актин-зв'язуючих білків, які специфічно взаємодіють з G-актином та F-актином. Ці білки виконують різні функції: регулюють обсяг G-актинового пулу (профілін), стабілізують кінці ниток F-актину (фрагін), зшивають філаменти з іншими компонентами цитоскелета. Деякі актин-зв'язуючі білки, наприклад, гельформуючі (від слова – желе) – скріплюють волокна актину хрест-навхрест і, тим самим, переводять стан ділянки цитоплазми зі стану золь (від латів. solutio – розчин) в гель. Ще один актин-зв'язуючий білок - спектрин, званий також фодрин, з'єднує волокна актина в пучки і прикріплює їх до цитоплазматичної мембрани і до сітки, побудованої з проміжних волокон. Білок валін зчеплює актинові філаменти в паралельно впорядковані жорсткі структури та впливає на швидкість полімеризації G-актину.
Майже всі типи рухів у клітині відбуваються за участю актин-зв'язуючого білка міозину. У всіх молекул міозину є головка, шийка та хвіст. Головка міозину здатна приєднуватися до мономеру актину і, за наявності АТФ, рухатися від плюс-до мінус-кінця мікрофіламенту. У скелетних м'язах молекули актину та міозину розташовані на фіксованих відстанях один від одного, а переміщення головок міозину по актинових нитках призводить до скорочення м'язів. У нем'язових клітинах при взаємодії з білком міозином актинові філаменти можуть формувати скорочувальні пучки, завдяки яким утворюються інвагінації (вп'ячування) клітинної поверхні. Такі інвагінації утворюються, наприклад, при розподілі клітин. Загалом характер рухів у клітині залежить від будови білка міозину, структура якого має більше 80 варіантів. Комбінуючи актинові мікрофіламенти з різними варіантами міозину та іншими актин-зв'язуючими білками, клітина формує структури, що відрізняються за архітектурою, рухливістю та часом існування.
Більшість клітин мікрофіламенти утворюють під плазматичної мембраною тривимірну структуру, так звану актинову кору (актиновий кортекс). Особливістю цієї структури є швидке оновлення мікрофіламентів; наприклад, у кортексі лейкоциту філаменти існують не більше 5 секунд. Основний тип перебудов кортексу у рухомих клітин пов'язаний із утворенням
псевдоподій – виростів цитоплазми. Псевдоподії можуть мати форму плоскої пластинки (ламелоподію), вузького циліндра (філоподію) або кулястого міхура. Форма псевдоподій залежить від типу актин-зв'язуючих білків, що взаємодіють з мікрофіламентами та плазматичною мембраною.
Актинові мікрофіламенти беруть участь у створенні складних просторових і щодо стабільних цитоскелетних структур. Наприклад, основу мікроворсинок епітеліальних клітин кишечника та нирок становлять довгі пучки актинових філаментів. На верхній поверхні волоскових клітин равлики внутрішнього вуха, які відповідають сприйняття звуків, перебувають спеціалізовані відростки (волоски) – стереоцилії. Стереоцилії розташовуються правильними рядами подібно до труб клавішно-музичного інструменту – органу. Внутрішня порожнина волоска-стереоцилії заповнена актиновими філаментами та молекулами інших білків. Мутації деяких генів, що кодують ці білки, призводить до дегенерації волоскових клітин та проявляється у вигляді однієї з форм спадкової глухоти (синдром Ашера).
Мікрофіламенти беруть активну участь у русі клітини. При цьому актинові філаменти постійно полімеризується на кінці рухового краю клітини та деполімеризується з внутрішньої сторони. Процеси полімеризації та деполімеризації F-актину можуть бути порушені отрутами (токсинами) грибів. Наприклад, фалоїдин (отрута блідої поганки) зв'язується з мінус-кінцем актину та інгібує деполімеризацію, у той час як цитохалазин (токсин з пліснявих грибів, що володіє властивістю цитостатика) приєднується до плюс-кінця, блокуючи полімеризацію актину та рух клітини. Тривалий вплив речовин, що порушують полімеризацію або деполімеризацію актинових філаментів, призводить до смерті цих клітин.
Полімеризація актину – це точно регульований процес, контрольований за допомогою поверхневих рецепторів клітини, ферментів (протеїнкіназ) та іонів кальцію. Порушення цього процесу супроводжується клінічними проявами. Наприклад, у трансформованих клітинах відзначається зменшення експресії білків, що регулюють складання актину. Значні аномалії актинових філаментів спостерігаються у клітинах деяких злоякісних пухлин. У клітинах саркоми (пухлини сполучної тканини) виявлено наявність тонких та коротких філаментів актину. Ці клітини, на відміну від нормальних клітин, дуже рухливі і мають велику здатність до метастазування.
Проміжні філаментискладаються з білків специфічних для певних клітинних типів (напр. кератини в епітеліальних клітинах, віментин у клітинах сполучної тканини, десмін у клітинах м'язових тканин та ін). Проміжні філаменти надають міцність клітині, так як вони є міцними, волокнистими, стійкими до розтягування поліпептиди і розподіляються по всій цитоплазмі клітини, утворюючи міцну мережу. Крім того, проміжні волокна присутні в ядрі, утворюючи мережу філаментів (ламіну) на внутрішній поверхні ядерної мембрани, що тісно пов'язану з ядерними порами.
Структурними елементами проміжних волокон є білки, що належать до п'яти родинних сімейств і виявляють високий рівень клітинної специфічності. Типовими представниками цих білків є цитокератини, десмін, віментин, кислий фібрилярний гліапротеїн та нейрофіламент. Всі ці білки мають у центральній частині базову стрижневу структуру, яка зветься α-спіралі. Два пептидні ланцюги (димери) утворюють суперспіраль. Такі димери з'єднуються антипаралельно, утворюючи тетрамер. Агрегація тетрамерів за принципом "голова до голови" дає протофіламент. Вісім протофіламентів сплітаються разом та утворюють проміжне волокно діаметром 10 нм. Еластичність проміжних філаментів забезпечується тим, що димери кожного тетрамера розташовані у шаховому порядку щодо один одного.
Волосся і нігті людини, шерсть, пір'я, голки, пазурі, і копита тварин складаються головним чином з кератину (цитокератину). В одному волокні вовни переплетені мільйони фібрил. Окремі ланцюги кератину скріплені численними дисульфідними зв'язками, що надає їм додаткової твердості. Виділено понад 30 різних кератинів, що комбінуються по два типи в епітеліальних клітинах людини. Крім того, описано вісім ізоформ важких кератинів, специфічних для волосся і нігтів. У нервових клітинах існують нейрофіламенти, що надають необхідної механічної опори довгим аксонам. Філаменти десміну розташовані в Z-дисках саркомерів скелетних м'язів. У різних типах клітин проміжні філаменти відіграють важливу роль у формуванні клітинних контактів, які називаються десмосомами, які з'єднують сусідні клітини. Напівдесмосоми прикріплюють епітеліальні клітини до базальної мембрани, де вони розташовані.
Мікротрубочки
Присутні у всіх еукаріотичних клітинах мікротрубочки є довгі ниткоподібні структури, протягнуті по всій цитоплазмі і формують мережу, яка підтримує структурну організацію і локалізацію деяких органел.
Мікротрубочки утворюються при полімеризації білка тубуліна (лат. tubula – трубочка), який є гетеродимером, утвореним субодиницями α- і β- тубуліна. У процесі полімеризації α -Тубулін одного димера контактує з β -Тубулін наступного димера з утворенням протофіламентів. Тринадцять тубулінових поздовжніх рядів протофіламентів (ниток), що йдуть по спіралі, утворюють мікротрубочку діаметром 24 нм та довжиною кілька мікрометрів.
Рис. Схема будови мікротрубочки, що показує як тубулінові поліпептиди, зв'язуючись один з одним, утворюють циліндричну стінку. А. Поперечний зріз Б - Короткий відрізок мікротрубочки.
Мікротрубочки здатні утворювати синглет, дублеті триплет.
Aмікротрубочка дублету або триплету складається з 13 протофіламентів.
Трубочки Bі Cскладаються з меншої кількості протофіламентів, зазвичай 10.

Полімеризація мікротрубочок відбувається в напрямку від голови до хвоста таким чином, що мікротрубочка має певну полярність: її кінці позначаються відповідно як плюс-і мінус-кінці. Мікротрубочки у клітині нестабільні. Вони можуть швидко збиратися та розбиратися. У клітці мінус-кінці пов'язані з центром організації мікротрубочок (ЦОМТ) - структурою, розташованої біля ядра, яка містить у тварин клітинах пару маленьких тілець - центріолей, утворених з мікротрубочок, що злилися. Як правило, мікротрубочки асоційовані з іншими білками (міозин, дінеїн, кінезин), які пов'язують мікротрубочки з іншими елементами цитоскелета та органелами. Кінезін забезпечує транспорт органел і везикул (бульбашок) з однієї частини клітини в іншу від плюс-кінця мікротрубочки до мінус-кінця, а дінеїн від мінус-кінця до плюс-кінця.
Відомі хімічні сполуки, здатні блокувати складання мікротрубочок (колхіцин, вінбластин) і стимулюють утворення стабільних мікротрубочок (таксол). Слід зазначити, що в організмі, що розвивається, можуть скластися умови (недолік кисню, знижена або підвищена температура і т.д.), коли порушується складання мікротрубочок у клітинах, це може спричиняти відхилення від нормального розвитку.
Мікротрубочки, як і актинові філаменти, беруть участь у підтримці форми клітини. Поряд із статичною функцією, мікротрубочки беруть участь у багатьох процесах, що протікають у всіх еукаріотичних клітинах: мейозі, мітозі, клітинному русі та секреції. Вони є напрямними «рейками» для транспорту органел. Разом з асоційованими білками мікротрубочки здатні здійснювати механічну роботу, наприклад, транспорт мітохондрій, переміщення синаптичних бульбашок, рух вій (волосоподібних виростів клітин в епітелії легень, кишечника та яйцеводів) та биття джгутика сперматозоїда. Пухирці, що утворюються апаратом Гольджі, прямують у різні місця клітини по мікротрубочкам строго за призначенням. Крім того, мікротрубочки у формі мітотичного веретена – найважливіша частина апарату, що забезпечує правильний розподіл хромосом між дочірніми клітинами при розподілі еукаріотичних клітин.
Функції мікротрубочок: 1) забезпечення розбіжності хромосом при розподілі клітин; 2) підтримка форми клітини; 3) участь у транспорті макромолекул та органел; 4) забезпечення рухливості джгутиків, вій.
Функції цитоскелета
Цитоскелет виконує три основні функції.
1. Служить клітині механічним каркасом, який надає клітині типової форми і забезпечує зв'язок між мембранною і органелами. Каркас є динамічною структурою, яка постійно оновлюється в міру зміни зовнішніх умов і стану клітини.
2. Діє як "мотор" для клітинного руху. Двигуни (скоротливі) білки містяться у м'язових клітинах, а й у інших тканинах. Компоненти цитоскелета визначають напрямок та координують рух, розподіл, зміну форми клітин у процесі зростання, переміщення органел, рух цитоплазми.
3. Служить як «рейки» для транспорту органел та інших великих комплексів усередині клітини.
Мікрофіламенти та проміжні волокна.
Мікрофіламенти, побудовані з F-актину, пронизують мікроворсинки, утворюючи вузли. Ці мікроволокна утримуються разом з допомогою білків, що найбільше важливі з яких є фімбрин і віллін. Кальмодулін та міозиноподібна АТФ – аза з'єднують крайні мікроволокна з плазматичною мембраною. .
Клітина може змінювати набір синтезованих білків цитоскелета залежно від умов, але цей процес повільний. Конструкція цитоскелета здатна швидко змінюватись навіть без синтезу нових молекул, за рахунок полімеризації та деполімеризації ниток. У клітині постійно йде обмін між нитками і розчином білків-мономерів у цитоплазмі. У багатьох клітинах приблизно половина молекул актину і тубуліну знаходиться у вигляді мономерів у цитоплазмі та половина входить до складу ниток мікрофіламентів. Клітина регулює стабільність ниток цитоскелета, приєднуючи до них спеціальні білки, що змінюють швидкість полімеризації. Загальний принцип функціонування цитоскелету - динамічна нестабільність. Наприклад, форму еритроциту як двояковогнутого диска підтримує примембранный цитоскелет з волокон, утворених білком спектрином. Спектрин пов'язаний з білком анкерином (anchor – якір), який з'єднується з білком цитоплазматичної мембрани, відповідальним за транспортування аніонів (Cl - , HCO - 3). Дефекти білків спектрину та анкірину викликають незвичайну форму еритроцитів. Такі еритроцити дуже швидко руйнуються у селезінці. Хвороби, які викликаються такими порушеннями, називають спадковим сфероцитозом або спадковим еліптоцитозом.

Рис. Цитоскелет еукаріотів. Актинові мікрофіламенти пофарбовані в червоний, мікротрубочки - зелений, ядра клітин - блакитний колір.

Цитоскелет утворюється трьома компонентами: мікротрубочками, мікрофіламентами та проміжними філаментами.
Мікротрубочкипронизують усю цитоплазму клітини. Кожна з них є порожнистим циліндром діаметром 20 – 30 нм. Стінка мікротрубочок утворена 13-ма нитками (протофіламентами), скрученими по спіралі одна над одною. Кожна нитка, своєю чергою, складається з димерів білка тубуліна. Синтез тубулінів відбувається на мембранах гранулярної ЕПС, а складання в спіраль – у клітинному центрі.
Відповідно, багато мікротрубочків мають радіальний напрямок по відношенню до центріолів. Звідси вони розповсюджуються по всій цитоплазмі.
Більшість мікротрубочок має закріплений («-») і вільний («+») кінці. Вільний кінець забезпечує подовження та укорочення трубочок. віях, а також центроміри хромосом. Якщо повністю зруйнувати мікротрубочки цитоплазми, вони відростають від клітинного центру зі швидкістю 1 мкм/мин. Руйнування мікротрубочок призводить до зміни форми клітини(Тваринна клітина набуває зазвичай сферичну форму). При цьому порушуються структура клітини та розподіл органел.
В клітці мікротрубочки можуть розташовуватися:
у вигляді окремих елементів;
у пучках, в яких вони пов'язані один з одним поперечними містками (відростки нейронів);
Ø у складі пар або дублетів (осьова нитка вій і джгутиків);
Ø у складі триплетів (центріолі та базальні тільця).
У двох останніх варіантах мікротрубочки частково зливаються один з одним.
Функції мікротрубочок:
а) підтримання форми та полярності клітини;
б) забезпечення упорядкованості розташування компонентів клітини;
в) участь у освіті інших, складніших органел (центріолі, вії тощо.);
г) участь у внутрішньоклітинному транспорті;
д) забезпечення руху хромосом при мітотичному розподілі клітини;
е) забезпечення руху вій.
Мікрофіламенти. Мікрофіламентами названі тонкі нитки білкові діаметром5 – 7 нм,що зустрічаються практично у всіх типах клітин. Вони можуть розташовуватися в цитоплазмі пучками, сіткоподібними шарами або поодинці.
Основним білком мікрофіламентів є актин, частку якого припадає до 5% від загальної кількості білків. Крім нього, до складу мікрофіламентів можуть входити міозин, тропоміозин, а також кілька десятків актинзв'язуючих білків. Молекула актину зазвичай має вигляд двох спірально скручених ниток. Безпосередньо під плазмолемою розташовується кортикальна мережа, в якій мікрофіламенти переплетені між собою та з'єднані один з одним за допомогою спеціальних білків, наприклад, філаміну. Кортикальна мережа обумовлює плавність зміни форми клітин, поступово перебудовуючись за участю актин-розщеплюючих ферментів. Тим самим вона перешкоджає різкій та раптовій деформації клітини при механічних впливах. Окремі мікрофіламенти кортикальної мережі прикріплюються до інтегральних та трансмембранних білків плазмолеми, а також до так званих адгезійних сполук (фокальних контактів), які пов'язують клітину з компонентами міжклітинної речовини або з іншими клітинами. Мікрофіламенти більш стійкі до фізичних та хімічних впливів, ніж мікротрубочки.
Основні функції мікрофіламентів:
1) забезпечення певної жорсткості та пружності клітини за рахунок кортикальної мережі мікрофіламентів;
2) зміна консистенції цитозолю, у тому числі під час переходу золю в гель;
3) участь в ендоцитозі та екзоцитозі;
4) забезпечення рухливості нем'язових клітин (наприклад, нейтрофілів та макрофагів), в основі якої лежить зміна форми клітинної поверхні внаслідок регульованої полімеризації актину;
5) участь у скороченні м'язових клітин та волокон;
6) стабілізація локальних випинань плазматичної мембрани, що забезпечується пучками поперечно зшитих актинових філаментів (мікроворсинки, стереоцилії);
7) участь у формуванні міжклітинних сполук (перепережувальні демосоми та ін).
Проміжні філаменти є сплетені білковими нитками канати.завтовшки близько 10 нм.Такий показник зумовив відведення ним проміжного місця між мікротрубочками та мікрофіламентами. Проміжні філаменти утворюють тривимірні мережіу клітинах різних тканин тваринного організму. Вони оточують ядро і можуть бути в різних ділянках цитоплазми, утворюють міжклітинні сполуки (десмосоми і напівдесмосоми), розташовуються всередині відростків нервових клітин.
Основні функції проміжних філаментів:
1) структурна;
2) опорна;
3) функція розподілу органел у певних ділянках клітини.
Цитоскелет - сукупність ниткоподібних білкових структур - мікротрубочок та мікрофіламентів, що становлять опорно-рухову систему клітини. Цитоскелет мають тільки еукаріотичні клітини, в клітинах прокаріотів (бактерій) його немає, що є важливою відмінністю цих двох типів клітин. Цитоскелет надає клітині певної форми навіть за відсутності жорсткої клітинної стінки. Він організує рух органоїдів у цитоплазмі (т. зв. перебіг протоплазми), що лежить в основі амебоїдного руху. Цитоскелет легко перебудовується, забезпечуючи у разі потреби зміну форми клітин. Здатність клітин змінювати форму обумовлює переміщення клітинних пластів на ранніх стадіях зародкового розвитку. При розподілі клітини ( мітоз) цитоскелет «розбирається» (дисоціює), а в дочірніх клітинах знову відбувається його самоскладання.
Цитоскелет виконує три основні функції.
1. Служить клітині механічним каркасом, який надає клітині типової форми і забезпечує зв'язок між мембраною і органелами. Каркас є динамічною структурою, яка постійно оновлюється в міру зміни зовнішніх умов і стану клітини.
2. Діє як "мотор" для клітинного руху. Двигуни (скоротливі) білки містяться у м'язових клітинах, а й у інших тканинах. Компоненти цитоскелета визначають напрямок та координують рух, розподіл, зміну форми клітин у процесі зростання, переміщення органел, рух цитоплазми.
3. Служить як «рейки» для транспорту органел та інших великих комплексів усередині клітини.
24. Роль методу імуноцитохімії у вивчення цитоскелета. Особливості організації цитоскелета у м'язових клітинах.
Імуноцитохімічний аналіз – метод, що дозволяє проводити імунологічний аналіз цитологічного матеріалу в умовах збереження морфології клітин. ІЦХ – один з безлічі видів імунохімічного методу: імуноферментного, імунофлюоресцентного, радіоімунного тощо. Основою ІЦХ-методу є імунологічна реакція антигену та антитіла.
Цитоплазма еукаріотичних клітин пронизана тривимірною сіткою з білкових ниток (філаментів), яка називається цитоскелетом. Залежно від діаметра філаменти поділяються на три групи: мікрофіламенти (6-8 нм), проміжні волокна (близько 10 нм) та мікротрубочки (близько 25 нм). Всі ці волокна є полімерами, що складаються з субодиниць спеціальних глобулярних білків.
Мікрофіламенти (актинові нитки) складаються з актину – білка, найпоширенішого в еукаріотичних клітинах. Актин може існувати як мономер (G-актин, «глобулярний актин») або полімеру (F-актин, «фібрилярний актин»). G-актин - асиметричний глобулярний білок (42 кДа), що складається із двох доменів. У міру підвищення іонної сили G-актин оборотно агрегує, утворюючи скручений лінійний в спіраль полімер, F-актин. Молекула G-актину несе міцно пов'язану молекулу АТФ (АТР), яка під час переходу в F-актин, повільно гідролізується до АДФ (ADP), тобто. F-актин виявляє властивості АТФ-ази.
Б. Білки проміжних волокон
Структурними елементами проміжних волокон є білки, що належать до п'яти родинних сімейств і виявляють високий рівень клітинної специфічності. Типовими представниками цих білків є цитокератини, десмін, віментин, кислий фібрилярний гліапротеїн [КФГП (GFAP)] та нейрофіламент. Всі ці білки мають у центральній частині базову стрижневу структуру, яка зветься суперспіралізованою α-спіралі. Такі димери асоціюють антипаралельно, утворюючи тетрамер. Агрегація тетрамерів за принципом "голова до голови" дає протофіламент. Вісім протофіламентів утворюють проміжне волокно.
На відміну від мікрофіламентів та мікротрубочок вільні мономери проміжних волокон навряд чи зустрічаються у цитоплазмі. Їхня полімеризація веде до утворення стійких неполярних полімерних молекул.
В. Тубулін
Мікротрубочки побудовані з глобулярного білка тубуліна, що є димером α- і β-субодиниць. Тубулінові мономери пов'язують ГТФ (GTP), який повільно гідролізується та ГДФ (GTP). З мікротрубочками асоціюють два види білків: структурні білки лки-транслокатори.
Саме висловлювання про цитоскелет було вперше запропоновано Кольцова, видатним російським цитологом на початку ХХ століття, який і відкрив їх у 1920р. Елементи цитоскелета зустрічаються у всіх еукаріотичних клітинах, а ось аналоги цих структур є і у прокаріотів. Ступінь виразності елементів цитоскелета у різних клітинах різна. Наприклад, клітини епідермісу шкіри особливо багаті на проміжні філаменти. У м'язових волокон більше актинових мікрофіламентів, а мікротрубочки більше зустрічаються у відростках нервових клітин, пігментних клітин. Загальними властивостями елементів цитоскелета є те, що це білкові фібрилярні полімери, що не гілкуються, здатні до збільшення площі поверхні і руйнування. Така нестабільність елементів цитоскелету призводить до рухливості клітини. Наприклад, зміни їх форми. Деякі компоненти цитоскелета за участю спеціальних додаткових білків можуть стабілізуватися та утворювати складні фібрилярні ансамблі, граючи роль каркасу. При взаємодії з іншими спеціальними білками, що відносяться до моторних білків або транслокаторів, компоненти цитоскелета можуть брати участь у різноманітних клітинних рухах.
Цитоскелет поєднує три підсистеми. Вони різняться за складом, ультраструктурою, за функціональними властивостями. Це система мікрофіламентів (актин-міозин), система мікротрубочок (тубулін-дінеїн) та система проміжних філаментів (10-нм філаменти).
Мікрофіламентиутворюють пучки в цитоплазмі рухливих клітин тварин, утворюючи так само шар (під плазмалемою) кортикальний, а в рослинних клітинах і грибах розташовуються в шарах цитоплазми, що рухається. Основним білком мікрофіламент є білок актин. Це комплекс кількох білків. Кожен білок у цьому комплексі кодується своїм геном. Виділяють два види актину – мономерну форму (глобулярну форму) G-актин., що містить молекулу АТФ. При полімеризації G-актину утворюється тонка фібрила, завтовшки приблизно 8 мкм. Ця структура називається F-актином. Актинові мікрофіламенти полярні за своїми властивостями. Це динамічні структури, які можуть збиратися та розбиратися залежно від співвідношення глобулярного та фібрилярного актину.
Нестійка фібрилярна система у клітинах стабілізується величезною кількістю допоміжних білків, які взаємодіють із F-актином. так, наприклад, білок тропоміозин забезпечує взаємодію кількох ниток актину, надаючи їм жорсткості. Білки філамін та альфа-актинін утворюють поперечні зчіпки між нитками F-актину, що призводить до утворення складної тривимірної мережі. Ця мережа надає гелеподібного стану цитозолю. Інші додаткові білки можуть зв'язувати філаменти пучки. Наприклад, білок фімбрину. Крім того, існують білки, які взаємодіють з кінцями мікрофіламентів і запобігають руйнуванню. Взаємодія F-актину з усіма допоміжними білками регулює агрегатний стан мікрофіламентів, забезпечуючи їхнє пухке або, навпаки, тісне розташування. І забезпечує їхню взаємодію з іншими компонентами.
Особливу роль у взаємодії з актином грає білок міозин. Він не відноситься до допоміжних білків. Він є другим основним компонентом актинової системи.
Міозин – сімейство подібних білків. У всіх цих білків у структурі виділяють головну чи моторну частину, яка відповідає за АТФазну активність комплексу. Другий компонент міозинових білків – шийка, пов'язана з декількома регуляторними білками. І третій компонент – хвостова частина, яка специфічна для кожного виду міозину та визначає функцію цього білка.
Весь цей комплекс міозинів поділяють на три типи: міозин I, міозин II та міозин V.
Міозин I. Являє собою мономерну молекулу.
Міозини II та V – димери. Їхня ділянка хвостової частини утворює так звану альфа-надспіральну структуру. 2 молекули міозину II можуть взаємодіяти між собою та утворювати фібрилу.
Міозин I та V беруть участь у взаємодії цитоплазми та мембрани, наприклад, у транспорті везикул. Механізм взаємодії цих білків, основних білків системи мікрофіламентів, починається із взаємодії міозинової головки з актиновим філаментом, що призводить до згинання ділянки молекул міозину та подальшого переміщення.
За кожен цикл міозинова голівка переміщається у напрямку позитивного кінця актинового філаменту за рахунок гідролізу однієї молекули АТФ на 5 – 25 нм. Тобто відбувається односпрямоване ковзання філамента актину щодо молекул міозину. Ця модель дістала назву моделі Хакслі. Теорія ковзних молекул.
Поперечно-смугасті м'язові волокна є збільшеною моделлю мікрофіламенту. Міофібрили є ниткою товщиною 1-2 мкм з темними і світлими ділянками, що чергуються. Одиницею будови міофібрили є саркомірабо ділянка між двома Z-дисками або білками. Функції Z-дисків полягає у зв'язуванні сусідніх структур одна з одною. Самі Z-білки є скоротними структурами.
Розмір саркомерів у розслабленому стані варіює від 1,8 до 2,8 мкм. Уздовж саркомера розташовуються три ділянки протофібрилу. Тонкі, пов'язані з Z-диском, які є нитками актину. І товсті нитки, представлені молекулами міозину. Розташовуються товсті нитки як у проміжках між нитками актина.
Головки молекул міозину взаємодіють з нитками актину і виникають актин-міозинові комплекси в результаті взаємодії двох самостійних білків. Активність цих комплексів у багато разів більша за АТФазні активи одного білка міозину.
Скорочення міофібрил відбувається за рахунок зменшення відстані між Z-дисками. Тобто. Довжина саркомера скорочується приблизно на 20 відсотків. Механізм скорочення полягає у кооперативному укорочуванні всіх саркомерів за довжиною міофібрили. В основі скорочення лежить переміщення відносно один одного тонких і товстих ниток, при цьому товсті нитки міозину входять у проміжки між нитками актину, зближуючи Z-диска.
Яку функцію виконує система мікрофібрил у складі цитоскелета:
1) Утворення скорочувального апарату клітини, що забезпечує рухливість.
2) Формування скелетних структур, здатних до власного руху за рахунок процесу полімеризації та деполімеризації актину (G-актин та F-актин).
3) Механомеханічне переміщення у процесах ендо- та екзоцитозу та цитотомії (розподіл тіла клітини).
Друга опорно-скоротлива частина цитоскелета – тубулінова система або система мікротрубочок.Ця система мікротрубочок має багато спільного з вже розглянутою актин-міозинової системою. Схожа на неї, по-перше, здатністю до полімеризації та деполімеризації. По-друге, також має полярність білкових ниток. По-третє, це велика кількість допоміжних білків.
Основний білок цієї системи – тубулін. Тубулін є гетеродимером. Складається з двох частин – альфа та бета тубуліна. Ці субодиниці при асоціації утворюють власне білок тубулін.
У процесі полімеризації молекули тубуліна об'єднуються таким чином, що бета-субодиниця взаємодіє з альфа-субодиницею, а альфа-субодиниця взаємодіє з бета-субодиницею.
Такі молекули вишиковуються одна за одною в довгі нитки протофіламенти.
Одночасно з налаштуванням протофіламенту в довжину при полімеризації відбувається налаштовування в ширину. У шаховому порядку. Завширшки максимум до 13 протофіламентів. Поздовжні протофіламенти скручуються в порожнисту трубочку, в якій кожен мономер тубуліну характеризується лінійним розміром 5 нм. Зовнішній діаметр циліндра, що утворився, дорівнює приблизно 25 нм. Ось такі мікротрубочки, які вийшли в результаті полімеризації окремих молекул тубуліна в цитоплазмі, називаються одиночними мікротрубочками. Це динамічні структури. Динамічна нестабільність- Найємніша характеристика трубочки. Вони швидко розбираються та швидко збираються. Цей процес залежить від співвідношення в клітині молекул одиночних і організованих мікротрубочки.
При достатньої концентрації білка тубуліна полімеризація відбувається спонтанно і швидкість полімеризації завжди вище одному з кінців мікротрубочки, який і називається позитивним кінцем. При недостатній концентрації тубуліна мікротрубочки розбиратимуться з обох кінців. Розбирання мікротрубочок сприяє, по-перше, зниження температури, а по-друге, цей процес вимагає присутності іонів кальцію.
Виділяють кілька типів речовин, алколоїдів рослини, які визначають швидкість розбирання чи збирання молекул тубуліна. Найпоширеніший алколоід колхіцин. Ця речовина взаємодіє з окремими молекулами тубуліна і запобігає полімеризації. Середній час життя приблизно дорівнює п'яти хвилинам. Такий стан притаманний інтерфази. Окремі мікротрубочки на кінці, що росте, подовжуються зі швидкістю 4-7 мкм/хвилину, а потім досить швидко вкорочуються. 14-17 мкм/м. У клітинах, що діляться, мікротрубочки збираються в особливу структуру. Організуються в ахроматичний веретено поділу, що забезпечує процеси розподілу генетичного матеріалу між дочірніми клітинами. Час життя цих мікротрубочок у складі ахроматичного веретена всього 15-20 сек. Вважається, що нестабільність мікротрубочок пов'язана із затримкою гідролізу ГТФ. Однак, 20% мікротрубочок залишаються відносно стабільними протягом 20 годин у диференційованих клітинах. Пов'язана ця стабільність із модифікацією тубуліна.
Самі мікротрубочки не є скоротливими, проте вони є обов'язковими компонентами клітинних органел, що рухаються, таких як вії, джгутики, ахроматичне веретено поділу, як мікротрубочки цитоплазми, які обов'язкові для внутрішньоклітинного транспорту, процесів екзоцитозу, ендоцитозу і транспорту всіх видів.
Цитоплазматичні одиночні мікронеми, локалізуючись у гіалоплазмі, виконують дві функції – каркасну (скелетну) та рухову скелетну полягає у стабілізації форми клітини. При штучному розчиненні їхня клітина втрачає свою форму і прагне стати кулею. Створюючи внутрішньоклітинні організації, мікротрубочки є чинниками орієнтованого руху внутрішньоклітинних структур.
Двигуна роль мікротрубочок полягає в тому, що вони створюють впорядковану векторну систему руху. Позитивні кінці мікротрубочок спрямовані від центру клітини до периферії. А наявність цих позитивних та негативно спрямованих полярних кінців мікротрубочок з динеїнами створюють можливість перенесення в клітині компонентів від периферії до центру.
Мікротрубочки ростуть із центру організації мікротрубочок (ЦОМТ).
У цих центрах мікротрубочки починають своє зростання від спеціальних ділянок і зростання здійснюється полярно. Нарощується позитивний кінці мікротрубочок. Як ЦОМТ в клітинах тварин головним чином бере участь матрикс клітинних центрів або центросоми. Своїми негативними кінцями мікротрубочки звернені до ЦОМТ і в них відбувається заякорювання. Під цим розуміють взаємодію із спеціальними білками, що обмежують набір мікротрубочок. У клітинах вищих рослин полімеризація мікротрубочок відбувається по периферії клітинного ядра, від якого трубочки розходяться радіально.
У більшості випадків в інтерфазних клітинах тварин організму новоутворення та зростання мікротрубочок походить від спеціальної освіти.
1) Мікротрубочки формують організовані структури входячи до складу вій, центріолей і джгутиків, обумовлюючи рух вій і биття джгутиків.
2) Мікротрубочки організуються в нитки ахроматичного веретена поділу при розподілі клітини.
3) Здійснюють транспорт усередині клітини, переміщуючи мембранні, секреторні та транспортні білки та органоїди.
4) Є цитоскелетом клітини, забезпечуючи утримання форми.
Лекція: клітинний центр (центросома)
Центросоми або клітинний центр було виявлено у 1875 році Флемінгом. У 1876 – Бенеденом. Розташовуються у геометричному центрі клітини. Вони характерні для клітин тварин. Їх немає у вищих рослин, у нижчих грибів та деяких найпростіших. У клітинний центр входять дрібні щільні тільця центріолі, зазвичай у парі. Пара центріолей – диплосома. У цій парі центріолі орієнтовані перпендикулярно один до одного. Диплосома оточена світлішою цитоплазмою, від якої відходять радіально тонкі фібрили – центросфера.
Основу будівлі центріолі складають розташовані по колу дев'ять триплетів мікротрубочок. Утворений дев'ятьма триплетами порожнистий циліндр має ширину приблизно 0,15 мкм, а довжину 0,3 – 0,5 мкм. Перша мікротрубочка триплета називається а-мікротрубочка. Вона повна мікротрубочка. Друга та третя мікротрубочки є не повними. Вони містять 11 протофіламентів і впритул примикають один до одного Іншими словами, ділянка, що з'єднує мікротрубочки є загальною.
Кожен триплет розташований приблизно під кутом 40 градусів до радіусу циліндра. Мікротрубочки складаються з тубуліна. Крім тубуліна до складу центріолі входять додаткові структури, представлені білком динеїном.
Зазвичай, в інтерфазних клітинах у складі диплосоми виділяють материнську центріоль та дочірню. Дочірня розташовується перпендикулярно до поздовжньої осі материнської центріолі. У центральній частині центріолі розташовується так звана втулка, представлена білком нексином. Нексин формує вирости, які називаються спицями, дев'ять спиць у напрямі кожного триплету. Об'єм, який займає всередині центріолі втулка зі спицями, може займати від 3\4 до 1\5. Поряд з диплосомою від материнської центріолі розташовуються у вигляді аморфного матеріалу вирости, які називаються придатками або сателітами материнської центріолі. У дочірньої придатків ніколи немає.
Систему мікротрубочок центріолей описують формулою 9+0. Навколо центріолей тонковолокнистий матрикс – муфта, в яку занурені мікротрубочки. У муфті є супутники (= перецентріолярні сателіти). Вони складаються з фібрилярних структур із трикутною ніжкою. Ніжка несе голівку. Контактують із дрібними тільцями. Сателіти - центри, на яких відбувається складання мікротрубочок.
Така морфологія диплосоми не є даною. Усе це дуже пластичною структурою. Будова та активність центросоми кардинально змінюється залежно від періоду клітинного циклу.
Клітинним цикломназивається час від початку утворення клітини до її власного поділу.
Періоди: розподіл (розподіл ядра і розподіл цитоплазми), становить приблизно 1\7 частина клітинного циклу. А решта – період підготовки до поділу (інтерфазу).
Для кожної стадії клітинного циклу характерні свої особливості метаболізму та морфології.
Під час поділу у клітинах знаходиться 2 центросоми. Клітина має 4 центріолі, вони розташовуються на полюсах клітини як 2 диплосом. Материнська центріоля на всіх стадіях мітозу оточена досить широкою зоною, шириною приблизно 0,3 мкм, представленою тонкими фібрилами. Ця зона називається центріолярним фібрилярним гало. Від цього гало радіально відходять мікротрубочки. Важливо, що дочірня центріоль не має ні гало, ні мікротрубочок. І така диплосома виконує функції формування веретена мітотичного апарату. Ахроматичне веретено поділу.
Зона диплосом, центросфера диплосом, звана періцентріолярним матриксом, є центром організації або полімеризації мікротрубочок (ЦОМТ). Це перша форма активності центріолей.
Центріолі – центри полімеризації мікротрубочок. До кінця телофази, коли практично відбулося поділ цитоплазми клітин, хромосоми починають деконденсуватись і утворюються нові дочірні ядра. Відбувається руйнування ахроматичного веретена поділу та трубочки веретена деполімеризуються. Клітинні центри теж змінюють свою структуру, саме материнська і дочірня центріолі втрачають взаємне перпендикулярне розташування і відходять друг від друга. Відстань варіює до 2 мкм. Ці центріолі на початку G1-періоду формують сателіти, від яких радіально відходять мікротрубочки. Центріолі стають місцем формування цитоплазматичних мікротрубочок. У міру зростання мікротрубочок зв'язок з областю центріолей втрачається і мікротрубочки вільно існують у цитоплазмі деякий час. І в клітині відбувається ніби конвейєрна зміна та репродукція цитоплазматичних мікротрубочок. Якщо заборонити клітині переходити в наступну фазу, то буде стадія спокою (G0-період).
Перехід клітини в стадію виконання своїх функцій пов'язаний з функціонуванням клітинного центру як структури, що формує вію або виріст плазматичної мембрани, заповненої аксонемою. Аксонема – осьова нитка.
Аксонема складається з дев'яти дуплетів мікротрубочок, які відростають з центріолей, і також розташовуються по колу радіально і в кожному дуплеті виділяють повну та неповну мікротрубочку. Крім дуплетів мікротрубочок, для вії характерна наявність двох одиночних центральних мікротрубочок, які оточені додатковим білком нексином у формі осьового або центрального циліндра. (9+2). Центріолі виконують функцію базального тіла.
При настанні періоду S клітинний центр виконує ще одну форму активності, а саме подвоєння числа центріолей. Розмноження центріолей не пов'язане з їх розподілом, а відбувається шляхом утворення зачатку або процентріолі, яка формується на стінці наявної центріолі перпендикулярно до кожної центріолі. Спочатку закладається дев'ять одиночних мікротрубочок, потім вони перетворюються на дев'ять дуплетів і потім у дев'ять триплетів. Таке нарощування називається дуплікація. Завдяки такому зростанню структур спочатку утворюється коротка дочірня центріоль, яка потім доростає до розмірів материнської. У S-періоді, одночасно з дуплікацією, материнська центріоля продовжує утворювати цитоплазматичні мікротрубочки.
Внаслідок процесу дуплікації біля кожної центріолі виростає нова центріоля. Дуплікація центріолей є пусковим механізмом або сигналом реплікації молекули ДНК. Після завершення S-періоду у клітці перебуває вже дві диплосоми.
Після наступає наступний період клітинного циклу. Постсинтетичний період, що прямо передує розподілу. Саме тоді зникають сателіти на материнській диплосомі. Обидві материнські центріолі покриваються фібрилярним гало і починають формувати тепер мітотичні мікротрубочки.
Крім цього, в цитоплазмі відбувається розпад мікротрубочок і клітина прагне набути кулястої форми. Клітини, які здатні до тривалого розмноження, вони повторюють ці події від циклу до циклу. Якщо ж клітина перебуває у стані G 0 періоду, то центріоль братиме участь, по-перше, у процесі полімеризації цитоплазматичних мікротрубочок, а по-друге, утворення апарату руху вій і трубочок.
Віїподіляються на дві групи: кінетоцилії, які характерні для спеціальних епітеліїв або вільно плаваючих клітин та первинні вії.
Вія являє собою тонкий циліндричний виріст у цитоплазмі з постійним діаметром 300 нм. Виріст від основи до верхівки покритий плазмолемою. Усередині виросту розташована структура аксонема, що складається в основному з тубуліна та динеїну.
Нижня проксимальна частина вії занурена в цитоплазму і називається базальне тільце. Діаметри аксонеми та базального тільця однакові. Аксонема у своєму складі має дев'ять дуплетів, що утворюють зовнішню стінку циліндра аксонеми. Дуплети мікротрубочок злегка повернені під кутом приблизно 10 градусів по відношенню до радіусу аксонеми. У дуплетах мікротрубочок так само розрізняють повну або А-мікротрубочку, що складається з 13 протофіламентів і В-мікротрубочку, неповну, вона має 11 протофіламентів. А-мікротрубочка несе на собі вирости, які спрямовані до В-мікротрубочки з сусіднього дуплета. Формують ці ручки додатковий білок динеїну. Денеїн представлений великими білковими комплексами, що складаються з 9 - 12 поліпептидних ланцюгів, що містять 2 - 3 глобулярні головки, пов'язані разом більш гнучкими лінійними ділянками. Кожна головка динеїну має активний центр взаємодії з молекулою АТФ. Від А-мікротрубочок до центру центрального циліндра відходять радіальні допоміжні білки, які формують спиці, що відходять від центрального циліндра.
Базальне тільце вії має таку ж будову, як і центріоль. Є ручки, втулка та спиці, розташовані в нижній частині базального тільця. На ділянці базального тільця, що примикає до плазмолеми, є дев'ять придатків, що йдуть від кожного триплету до плазматичної мембрани і зв'язують його з клітинною тканиною. Тому базальне тільце і вія структурно пов'язані один з одним і становлять єдине ціле. А- і В-мікротрубочки в триплетах базального тільця, продовжуються в А- і В-мікротрубочках в дуплетах аксонем. А ось внутрішні частини відмінні один від одного і часто в зоні переходу базального тільця в вію спостерігають аморфну поперечну пластинку, від якої починаються в область аксонеми зростання центральних мікротрубочок. Вії не скорочуються. Вони згинаються чи б'ються. У цьому русі дінеїн є мото-або руховим білком. При асоціації динеїну із субодиницями тубуліна відбувається поздовжнє ковзання дуплетів один щодо іншого. Відбувається переміщення головок мікротрубочок від позитивного кінця до негативного кільця та сусідній дуплет зсувається до верхівки вії. Дуплети мікротрубочок пов'язані один з одним допоміжними білками з центральною парою мікротрубочок. Таке кооперативне зміщення дуплетів у бік верхівки призводить не до подовження вії, а до вигину. Процес є енергозалежним.
Багато бактерій здатні до руху за допомогою інших органел. Це бактеріальний джгутик або флагела. Джгутики бактерій принципово відмінні. Вони мають складну будову. Складаються з трьох основних частин: зовнішньої довгої волокнистої нитки власне джгутика, гачочка та базального тільця. Джгутикова нитка побудована з білка флагеліну. Молекулярна маса його від 40 до 60 тис. глобулярні субодиниці флагеліну полімеризуються в спірально закручені нитки так, що утворюється структура. Діаметр 12 – 25 нм. Порожня всередині. Білки флагеліну не здатні до руху. Вони можуть спонтанно полімеризуватись у спіральні нитки з певним кроком спіралі.
Поблизу клітинної поверхні бактерій флагела переходить до ширшої ділянки, яка називається гачок.
Довжина гачка близько 45 нм. Він складається з інших білків.
Бактеріальне базальне тільце складається із стрижня пов'язаного з гачком та чотирьох кілець «дисків». Одне кільце занурене у ліпосахаридну мембрану, інше – у шар муреїну. Інші занурені у білковий комплекс. У еукаріотів джгутики рухаються за рахунок поздовжнього руху дуплетів. У бактерій рух джгутиків відбувається за рахунок обертання базального тільця навколо осі в площині плазматичної мембрани. Рух джгутиків залежить від АТФ.
Третя складова частина – 10-нм проміжні філаменти. Вони будуються із фібрилярних мономерів. Основна конструкція проміжних філаментів нагадує канат, що має фіксовану товщину.
Локалізація проміжних філаментів суворо центрована у клітині. Вони розташовуються в навколоядерній зоні та в пучках фібрил, що відходять до периферії клітин.
Трапляються проміжні філаменти у всіх типах клітин тварин, але особливо багато в тих клітинах, які схильні до механічних впливів. Наприклад, епідерміс, м'язи, нервові відростки. У клітинах рослин немає проміжних філаментів.
До складу філаментів входить велика група ізомерних білків, які поділяються на чотири групи:
1) Кератинові волокна. Вони здатні до полімеризації. Складаються із двох підтипів. Поділяються на кислі та нейтральні.
2) Віментин, Віментинові волокна, які характерні для мезенхімних тканин. Десмін. Характерний для м'язової тканини, причому, і гладкої та поперечно-смугастої. Гліальний білок – оболонка навколо нейронів.
3) Нейрофіламенти. Аксони нервових клітин.
4) Білки ламіни. Вони не знаходяться в субмембранному шарі клітини, але останні дані показали, що за будовою та властивостями ламіну є проміжними філаментами.
Для всіх проміжних білків характерна подібна амінокислотна послідовність, представлена 130 залишками амінокислот в центральній частині фібрили, мають спіральну будову - альфа-спіраль (однакова у всіх).
Кільцеві ділянки характеризуються різною амінокислотною поверхнею, різною довжиною, не представлені спіраллю.
Наявність центральних доменів дозволяє утворити подвійну спіраль – димер довжиною близько 48 нм. Димери асоціюють пліч-о-пліч. Утворюють короткий проторфіламент, у якому буде вже 4 первісні молекули і називається він тетрамер. Товщина близько 3 нм. Протофіламенти ще раз визначаються попарно і утворюються довгі тонкі фібрили з восьми поздовжніх протофіламентів (октамер, діаметр 10нм). У цьому вся особливість полімеризації проміжних філаментів.
Білки ядерної ламіни, вони полімеризуються інакше. Вони утворюють димери з головками одному кінці, вони, полиризуясь по 2, формують пухку сітчасту прямокутну решітку. Такі грати пов'язані з димерів, здатна до реакції фосфорилювання, що призводить до розпаду пухких прямокутних решіток. Цитоплазматичні проміжні філаменти відносяться до найстабільніших і довгоживущих елементів цитоскелета. Це і є істинно-опорна система.
Цікаво, що розташування проміжних філаментів дублює розташування мікротрубочок. При руйнуванні мікротрубочок спостерігається цікаве явище, яке називається колапс проміжних філаментів. Вони збираються у щільні пучки навколо ядра.
Функції проміжних філаментів:
1) Структурна, протидіє силам розтягування;
2) Інтеграція трьох систем клітини: поверхневого апарату, цитозолю та ядра.
Підсумок теми.У складі цитоскелета можемо виділити такі елементи цитоскелета: тільки каркасні (проміжні філаменти) та опорно-рухові (актин-міозин, тубулін-дінеїн). В опорно-рухових елементах існують 2 різні способи руху:
1) Заснований на здатності основного білка мікрофіламентів актину та основного білка мікротрубочок тубуліна до полімеризації та деполімеризації, що призводить при зв'язку цих білків з плазматичною мембраною до її морфологічних змін у вигляді утворення псевдоподій, з метою переміщення клітин на поверхню субстрату.
2) При другому способі пересування фібрили актину або тубуліна є напрямними структурами, якими переміщуються спеціальні рухливі білки - мотори. Вони взаємодіють із мембранними чи фібрилярними компонентами клітини, викликаючи її переміщення.
-Сукупність ниткоподібних білкових структур - мікротрубочок і мікрофіламентів, що становлять опорно-рухову систему клітини.
Цитоскелет – високодинамічна система цитоплазми. Багато структур цитоскелета можуть легко руйнуватися і знову виникати, змінюючи своє розташування або морфологію. В основі цих особливостей цитоскелета лежать реакції полімеризації-деполімеризації основних структурних цитоскелетних білків та їх взаємодія з іншими білками як структурними, так і регуляторними.
Цитоскелет мають тільки еукаріотичні клітини, в клітинах прокаріотів (бактерій) його немає, що є важливою відмінністю цих двох типів клітин. Цитоскелет надає клітині певної форми навіть за відсутності жорсткої клітинної стінки. Він організує рух органоїдів у цитоплазмі (т. зв. перебіг протоплазми), що лежить в основі амебоїдного руху. Цитоскелет легко перебудовується, забезпечуючи у разі потреби зміну форми клітин. Здатність клітин змінювати форму зумовлює переміщення клітинних пластів ранніх стадіях зародкового розвитку. При розподілі клітини (мітозі) цитоскелет «розбирається» (дисоціює), а в дочірніх клітинах знову відбувається його самоскладання.
Функції цитоскелета різноманітні. Він сприяє підтримці форми клітини, здійснює всі типи клітинних рухів. Крім того, цитоскелет може брати участь у регуляції метаболічної активності клітини.
Цитоскелет утворений білками. У цитоскелеті виділяють кілька основних систем, званих або за основними структурними елементами, помітними при електронно-мікроскопічних дослідженнях (Мікрофіламенти, проміжні філаменти, мікротрубочки), або за основними білками, що входять до їх складу (актин-міозинова система, кератини, тубулін-дінеїн) ).
Проміжні філаментиє найменш зрозумілою структурою серед основних компонентів цитоскелета щодо їх складання, динаміки та функцій. Їх властивості та динаміка сильно відрізняються від відповідних характеристик як мікротрубочок, так і актинових філаментів. Функції ж проміжних філаментів досі залишаються у сфері гіпотез.
Цитоплазматичні проміжні філаменти виявлені у переважній більшості укаріотичних клітин, як у хребетних, так і безхребетних тварин, у вищих рослин. Рідкісні приклади клітин тварин, які не виявлені проміжні філаменти, що неспроможні вважатися остаточними, оскільки білки проміжних філаментів можуть утворювати незвичайні структури.
Морфологічні мікротрубочкиє порожнистими циліндрами діаметром близько 25 нм з товщиною стінки близько 5 нм. Стінка циліндра складається з протофіламентів - лінійних полімерів тубуліна з поздовжньо орієнтованими гетеродимерами. У складі мікротрубочок протофіламенти йдуть уздовж їх довгої осі з невеликим зрушенням один щодо одного, так що субодиниці тубуліна утворюють тристартову спіраль. До складу мікротрубочок більшості тварин входить 13 протофіламентів
Актинові філаментивідіграють ключову роль у скорочувальному апараті м'язових та нем'язових клітин, а також беруть участь у багатьох інших клітинних процесах, таких як рухливість, підтримання форми клітини, цитокінез
Актинові філаменти або фібрилярний актин (F-актин) є тонкими фібрилами діаметром 6-8 нм. Вони є результатом полімеризації глобулярного актину – G-актину. У клітині актинові філаменти з допомогою інших білків можуть утворювати безліч різноманітних структур.